In a globalized world, changes in human activities are negatively affecting biodiversity at different scales. Long-term monitoring of bird communities can inform about changes in biodiversity over time. We explored how bird species richness, and abundance changed after a 37-year period at 5 sites in O Courel Mountains, a well conserved region of NW Spain. We also analyzed the abundance changes over time in the 15 most abundant species and whether these changes relate to the phenotypes of these species. Repeated bird surveys were performed in the same locations of five different sites in 1977 and 2014. Our results showed species richness and overall bird abundance did not change after 37 years within the five studied sites. Although, within species analyses showed 8 species changed their abundances. Overall, Cyanistes caeruleus, Garrulus glandarius, Parus major, Prunella modularis and Sylvia atricapilla increased their abundances and Erithacus rubecula, Phylloscopus ibericus, and Troglodytes troglodytes decreased over time. We did not find any relationship between the change in species abundances and bird phenotypes. This study shows that, despite the specific changes in the abundance of eight species over nearly four decades, overall bird species richness and abundance may be maintained in well preserved natural areas.
Palabras clave:
bird diversity, long-term diversity changes, bird traits, well preserved areas
Losada, Salaverri, Docampo, Guitián, and Sobral: Bird communities after 37 years in a rural area of NW Spain
Introduction
Birds are excellent biodiversity indicators because their population trends are affected
by many ecological factors, such as the niche breadth () and the climatic niche (). They are very sensitive to environmental changes (; ; RUSSO, 2007), although sensitivity differs among species (; ). But also birds constitute a conspicuous and cosmopolite animal group generally
easy to sample (). Due to these reasons, birds are commonly used as model species in long-term monitoring
studies of diversity changes (; ; ) or population trends (; ).
Human-induced environmental changes are impacting biodiversity (; ; ; ), such as bird species richness and abundance (; ; ). For example, many western humanized areas have seen a gradual rural abandonment
of traditional agriculture throughout the 20th century (; ; ; ), while others have experienced the process of industrial agricultural intensification
(; ). As a consequence, in both cases, biodiversity levels might decrease as has already
been observed in plant communities in different regions of Europe (; ; ; ) but also in bird communities (; ; ; ; ; ; ).
Over the last 50 years, rural Europe, and within it, the NW of the Iberian Peninsula,
have undergone a strong process of rural abandonment. Within the natural region of
northwestern Spain, O Courel (42° 34′ 59.99″ N, 7° 10′ 0.01″ W; 21,020 ha), the number
of inhabitants was reduced (-60%) and with it, the livestock (-80%) and number of
farmlands (-36%) decreased as well (; ). All these changes have modified vegetation patterns favoring woodland areas (; ), with an associated loss of bird diversity () which may be linked to a landscape homogenization (; ).
However, within O Courel region, areas which have not been cultivated remained stable,
thus a vast majority of the region has not locally changed (). We do not know whether and how bird richness and abundance may have changed in
the well-preserved areas of this region after a 37-year period. Analyzing changes
in the richness and abundance of bird communities after several decades in well preserved
plots in NW Iberia would help us understanding bird diversity trends at undisturbed
sites, i.e. which have not undergone abandonment or industrialization.
We investigated changes in bird diversity and species trends after a long-term period
in different habitats which have remained well preserved within O Courel region. Here,
we assessed changes in overall bird species richness and abundance within five sites
(two chestnut woodlands, a montane forest, a holm oak forest and a heathland) after
37 years, using data obtained from bird surveys performed in the same locations with
the same methodology in July 1977 and July 2014. Bird populations might show different
trends over time, and this could be related to their specific traits (; ; ; ; ). Thus, we additionally analyzed whether species phenotypes are related to the abundance
trends of the 15 most abundant species overall in this area. We hypothesized that
(i) the overall number of species and bird abundance varied after a 37-year period,
(ii) and these changes may differ among sites, and (iii) changes in species abundances may be explained by bird phenotypic traits.
Materials and Methods
Study area
The study was performed in O Courel Mountains (Galicia, NW Spain), mostly located
within the municipality of Folgoso de Courel (Lugo: 42° 35' 20" N -7° 11' 43" W; 193
km2), and part of the EU Natura 2000 network included in the “Ancares-Courel” Site of
Community Importance (Nat-2000 Site Code ES1120001). Annual rainfall exceeds 1,500
mm, snow warnings are probable between November and April, and the annual average
temperature oscillates between 8 and 10ºC, although it can even reach 23ºC (). About 750 species of higher plants are present in this 21,020-ha region, as well
as, rare or very rare plant species in Galicia (). One of the most remarkable vegetal units are the heathlands covering 70% of the
total surface, the common hawthorn woodlands (Crataegus monogyna) over the limestone outcrops; the montane mixed forests mainly composed by Pyrenean
oaks (Quercus pyrenaica) and Holm oaks (Quercus rotundifolia Lam.) occupying 20% of the total area; but also the chestnuts woodlands (Castanea sativa). Furthermore, this region is home to 114 bird species (; ), and it is also important to mention the secular human presence in this area. The
most notable human land uses were croplands on the less steep slopes, logging, and
grazing activities in the scrubland areas ().
We selected 5 sites belonging to 4 habitats for this study (three different leaf deciduous
forests and a heathland), mostly located in the Moreda valley, with the exception
of the heathland in the Ferreirós valley: (1) Devesa da Rogueira (hereinafter montane
forest) is a montane mixed forest (900 - 1,400 m.a.s.l.) with a dense undergrowth
(Erica arborea and Vaccinium myrtillus) and a great tree diversity (Fagus sylvatica, Quercus petraea, Sorbus aucuparia and Taxus baccata; ). (2) Moreda’s holm oak forest (from now on, holm oak forest) is dominated by Quercus ilex subsp. rotundifolia L. on a limestone substrate (800 - 900 m.a.s.l.) and accompanying Quercus pyrenaica and Crataegus monogyna tree species. (3) De Lomba’s heathland (hereinafter, heathland) is mainly composed
of Erica australis and Genista tridentata, growing on skeletal soil (1,200 - 1,400 m.a.s.l.) and forming very low but dense
heaths due to consecutive fires. (4) Fuente Parada’s chestnut woodland and (5) Moreda’s
chestnut woodland (hereinafter) are located on a valley floor slope (approximately
900 m.a.s.l.), primarily composed of Castanea sativa trees (see Table SIa, for more site details in Supplementary materials section).
Bird surveys
This study used bird survey data collected on July 1977 () and July 2014, using an identical methodology in the five locations (Data S1 in Supplementary materials section). The transects were surveyed in 1977 (3-8 times) and in 2014 (twice) following the
same itinerary and walking the same distance (montane forest: 500 m; Holm oak forest:
530 m; heathland: 615 m; Parada’s chestnut woodland: 330 m; Moreda’s chestnut woodland:
360 m) with the exception of the holm oak forest since the original trail disappeared
and an alternative route (156 m longer) was performed in the same forest plot in 2014
(Table SIb in Supplementary materials section). During each survey, all the individuals observed and heard within a 25-m band on
each side of a finish transect were recorded maintaining a constant walking speed
(). All the surveys were performed by expert ornithologists (José Guitián in 1977 and
jointly Luis Salaverri in 2014) early in the morning and under windless and rainless
weather conditions.
Bird traits
Bird phenotypic data were extracted from the “Life-history characteristics of European birds” open-access dataset () and available at Dryad: https://datadryad.org/stash/dataset/doi:10.5061/dryad.n6k3n. We selected 27 numeric trait variables with complete cases for all the 15 most abundant
species in our study (Cyanistes caeruleus, Erithacus rubecula, Fringilla coelebs, Garrulus glandarius, Phylloscopus
ibericus, Parus major, Prunella modularis, Periparus ater, Regulus ignicapilla, Sylvia
atricapilla, Sitta europaea, Serinus serinus, Sylvia undata, Turdus merula, Troglodytes
troglodytes). The phenotypic traits included were the mean bird length (cm); the mean wing length
(mm), the mean tail length (mm), the mean bill length (mm), the mean tarsus length
(mm), the mean weight during breeding (g), and the corresponding means of all these
traits by sex. With regard to the reproductive traits, we considered the minimum,
the maximum and the mean of clutch size (no. of eggs), the mean brood size per year
(as the average no. of broods per breeding season), the mean length and mean width
of the eggs (mm), the mean egg mass (g), the incubation period (as the average egg
incubation period, days), the fledging period (as the average age of youngs when fledgling,
days), the feeding independence (as the average age of youngs when being independent
of parental feeding, days) and the age of first breeding (as the average age of first
breeding event, years).
Statistical analyses
Changes in the number of species between years and among sites, were analyzed by a
generalized linear model (GLM) where the response variable was the total number of
bird species observed per site and year (n= 10), fitted to a normal error distribution
with an identity-link function. Overall bird abundance differences between years and
among sites were also analyzed by a generalized linear model (GLM) where the response
variable was the sum of individuals recorded per site and year (n=10), corrected by
the number of surveys (see Table SIb in Supplementary materials section) fitted to a normal error distribution with an identity-link function. In both models,
year (1977 and 2014) and site (montane forest, holm oak forest, heathland, Parada’s
and Moreda’s chestnut woodlands) were included as fixed effects. This approach allows
us to analyze the global variation in the number of species and bird abundance between
sites and between years (the objective of our study), and not only the variability
of bird richness and abundance per site, in which case we would only have one annual
replicate per site, both for overall species richness and bird abundance. In addition,
number of species and birds per survey was low and not representative of each site
(Supplementary Data S1). Thus, we aggregated richness and abundance data as the survey sum of species and
individuals per site and year. This is why the interaction between site and year factors
was not included in both models.
Considering the 15 most abundant bird species (i.e.10 or more individuals recorded;
Table II, Fig. 1), we analyzed the abundance change between 1977-2014 and among the five study sites
by constructing an independent generalized linear model (GLM) for each species, in
which the response variable was the sum of individuals observed per site and year,
corrected by the number of surveys (Table SIb in Supplementary materials section), fitted to a normal error distribution with an identity-link function and including
the year and site as fixed effects.
For the analysis of the relationship between abundance changes and bird phenotypes,
we first created a variable at the species level with three categories for the abundance
change factor accordingly to the sign of the estimated year effect (“+”: species with
statistically significant increase in abundance, “-”: species with statistically significant
decrease in abundance, “ns”: species with non-significant change in abundance) based
on the results of the per species abundance analysis explained above. Second, we selected
27 numeric avian phenotypic traits from Dryad open-source dataset (; see above bird traits sub-section for more details) to perform a Principal Component
Analysis (PCA) for dimensional reduction. We extracted three Principal Components,
with initial eigenvalues from the matrix of variance above 1.0 and which explained
a total of 91.66% variance of 27 phenotypic traits among our 15 more abundant species
(PC1: 72.33%; PC2: 15.38%; PC3: 3.95%). PC factor loadings were included in the Supplementary
materials section (see Table SII).
Change in abundance per species as coded above (increase, decrease and non-significant
change) in function of bird phenotypic traits was analyzed by means of a generalized
linear model (GLM) in which the response variable was the factorial abundance change
(three levels: increase in five species, decrease in three species, non-significant
change in seven species) per species (n= 15), fitted to a multinomial error distribution
with a log-accumulated link function (the total change in species abundance was analyzed
in a similar manner and gave equivalent results, not shown). The scores of three Principal
Components extracted from PCA were included as explanatory covariates (analyses of
individual phenotypic traits were additionally tested and gave equivalent results,
not shown).
All analyses were performed using the SPSS for Windows, version 25.0 (IBM CORP. 2017).
Error distribution, link function and model's structure were chosen by means of the
AICc criterion starting with saturated models ().
Results
Overall, 392 individuals of 31 bird species were recorded.
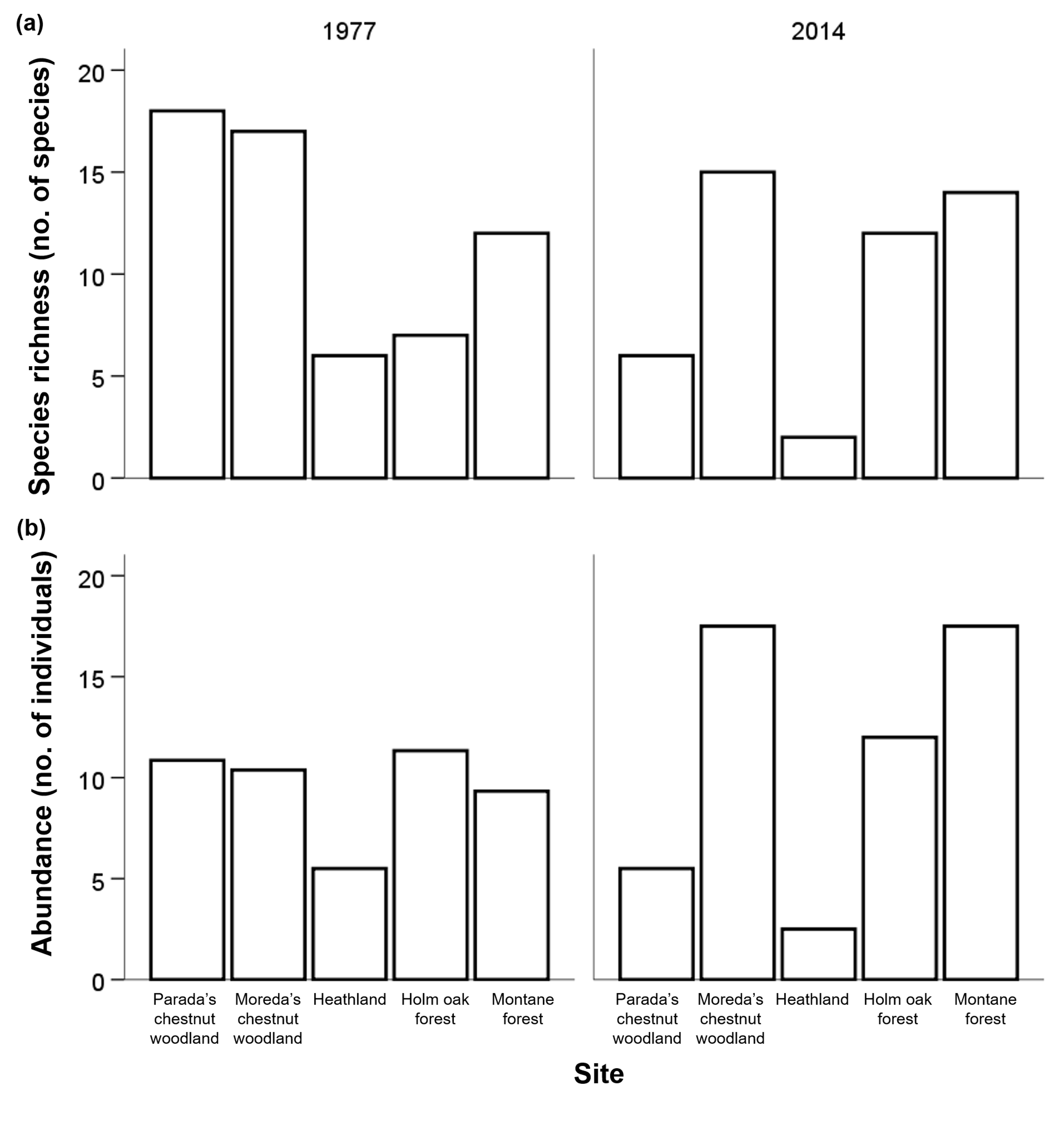
The number of bird species varied among sites but did not change overall from 1977
to 2014 (Table I, Fig. S1a). The heathland was the site with the lowest specific richness (6 species in total
for both years). Similarly, the overall abundance of birds (sum of individuals per
site and year corrected by the number of surveys) has not changed after the 37-year
period although it differed among sites (Table I, Fig. S1b), being the heathland the site with lower bird abundance (8 birds in total for both
years).
Table IGLMs results for richness (total no. of species observed per site and year) and abundance
(sum of birds observed per site and year, corrected by the corresponding no. of surveys)
variation in function of the year (1977, 2014) and the site (Paradas’s chestnut woodland,
Moreda’s chestnut woodland, heathland, holm oak forest, and montane forest) factors
included as fixed effects into the models (n = 10). Marked in bold are the statistically
significant effects.
Response variable
Fixed effects
Wald Chi-square
d.f.
P-value
Richness (no. of species)
Year
1.434
1
0.231
Site
19.242
4
0.001
Abundance (no. of individuals)
Year
0.802
1
0.370
Site
19.156
4
0.001
Among the 15 more abundant species (i.e. more than 10 individuals observed in both
years), only eight species showed a change in their abundance over time and 13 species
differed in their abundance among sites (Table II). In particular, the abundance of Cyanistes caeruleus, Garrulus glandarius, Parus major, Prunella modularis and Sylvia atricapilla increased between 1977-2014, and Erithacus rubecula, Phylloscopus ibericus, and Troglodytes troglodytes decreased in their abundances after the same 37-year period (Table II, Fig. 1). Among the seven species that did not differ in abundance between years: Fringilla coelebs, Periparus ater, Regulus ignicapilla, Sylvia undata and Turdus merula showed differences in the number of individuals detected per species among sites,
as did the 8 species that varied in abundance over time (Table II, Fig. 1).
Table IIGLM results for species abundance (sum of individuals observed per site and year,
corrected by the no. of surveys) variation in function of the year (1977, 2014) and
the site (Paradas’s chestnut woodland, Moreda’s chestnut woodland, heathland, holm
oak forest, and montane forest) included as fixed effects in the independent models
performed for the 15 most abundant species, i.e. 10 or more individuals (n = 10).
We marked in bold the statistically significant effects, and specified the sign of
the significant changes between years (“+”: increase; “-”: decrease, “ns”: non-significant
change).
Response variable
Species
Fixed effects
Wald Chi-square
d.f.
P-value
Change
Abundance (no. of individuals)
C. caeruleus
Year
5.330
1
0.021
+
Site
26.408
4
<0.001
E. rubecula
Year
5.358
1
0.021
-
Site
225.432
4
<0.001
F. coelebs
Year
0.357
1
0.550
ns
Site
14.988
4
0.005
G. glandarius
Year
24.323
1
<0.001
+
Site
223.990
4
<0.001
P. ibericus
Year
6.265
1
0.012
-
Site
10.000
4
0.040
P. major
Year
18.045
1
<0.001
+
Site
13.068
4
0.011
P. modularis
Year
10.808
1
0.001
+
Site
31.557
4
<0.001
P. ater
Year
0.908
1
0.341
ns
Site
14.571
4
0.006
R. ignicapilla
Year
3.623
1
0.057
ns
Site
24.371
4
<0.001
S. atricapilla
Year
6.552
1
0.010
+
Site
38.620
4
<0.001
S. europaea
Year
2.071
1
0.150
ns
Site
9.142
4
0.058
S. serinus
Year
0.000
1
0.986
ns
Site
8.349
4
0.080
S. undata
Year
2.500
1
0.114
ns
Site
20.408
4
<0.001
T. merula
Year
0.873
1
0.350
ns
Site
11.695
4
0.020
T. troglodytes
Year
12.59
1
<0.001
-
Site
13.59
4
0.009
Figure 1Change in species abundance (sum of individuals per site and year, corrected by the
number of surveys) of the 15 more abundant species (i.e. ≥ 10 individuals observed
overall) by site: montane forest (light green-colored bars), holm oak forest (yellow-colored
bars), heathland (dark green-colored bars), Moreda’s chestnut woodland (red brown-colored
bars) and Parada’s chestnut woodland (brown-colored bars).
With regard to bird traits analysis, we did not find any relationship between the
phenotypes of bird species (measured by PCA from the phenotypic space using 27 traits,
Table SII in Supplementary section) and the specific change in species abundance over 37 years
for the 15 more abundant species (Table SIII, Fig. 2).
Figure 2PC2 factor (15.38 % of variance explained) in function of PC1 factor (72.33 % of variance
explained) extracted from PCA for phenotypic traits variables reduction (n= 15 species),
according to the factorial abundance change (green dots: increasing species, red dots:
decreasing species, blue dots: species with non-significant change).
Discussion
The overall number of bird species and individuals did not change after 37 years,
in well-preserved areas from NW Spain (O Courel mountains) but we found specific changes
in abundance of 8 of 15 most abundant species. These species specific trends may depend
on their characteristics, their preferences for a certain land cover, their migration
patterns or their biogeographical origin (). For example, species traits may determine their level of success in particular
habitats, where certain level of specialization is required for breeding and surviving
(; ; ). However, we did not find a relationship between species traits and abundance change
in 15 most abundant bird species after 37 years, maybe due to the lack of sufficient
phenotypic variation among these 15 species compared here.
We found differences in species richness and overall bird abundance among sites, being
the heathland the site with the lowest number of species and abundance compared to
the other sites. These differences observed among sites may be linked to differences
in floristic composition and vegetation structure (; ; ), as well as, land cover (). Thus, the higher number of species observed corresponds to mature deciduous forests
(i.e. with a more diverse plant community and complex vegetation structure): Moreda’s
(20 species in total) and Parada’s (19 species) chestnut woodlands, the montane forest
(17 species overall), and the holm oak forest (14 species in total), but the lowest
species numbers was found in the heathland (6 species overall), the site with lower
plant diversity and more simple vegetation structure. However, bird species numbers
found in our mature deciduous forests were rather lower than those found in other
NW Iberian forests (30-37 species; ; ; ), which may be explained by altitudinal differences with regard to our study sites,
that have already accounted for a decrease in bird species at higher elevations (), specially forest-dwelling species (), but also by the landscape structure, in particular when considering the surrounding
areas (). Thus, marked habitat differences among sites may explain the reduced number of
bird species and abundance found in the heathland.
Moreover, the greater numbers of birds (as the sum of individuals per site and year
corrected by the number of surveys) were found in the Moreda’s chestnut woodland (28
birds in total for both years), montane forest (27 birds in total), and the holm oak
forest (23 birds in total), and these numbers are rather lower than those observed
in the already mentioned NW Iberian forests (60-140 total birds; ; ; ). If we compare bird abundance among study sites, our results may suggest that mature
deciduous forests could be acting as bird diversity reservoirs due to a possible landscape
homogenization of the surrounding plots (), as a consequence of the ongoing farmland abandonment process (; ; ), but also the increase of woodland areas in this region (; ). These results may be supported by the fact that O Courel still maintains well preserved
areas given the landscape heterogeneity provided by a mosaic of large patches, mainly
dominated by leaf deciduous forests and scrublands ().
Therefore, the bird richness and abundance remained stable after 37 years at the study
sites in O Courel Mountains, despite the specific changes in species trends of 8 most
abundant species found within our study area. Although we were not able to analyze
the effect of the year*site interaction (given the level of our analysis), it seems
that these eight species specific trends might differ when we separate the change
in species abundance by site (Fig. 1). One possible explanation to the specific changes in species abundance is the habitat
preference of these species (). The forest-dwelling species (Cyanistes caeruleus, Garrulus glandarius, and Parus major) showed a generalized increase, which is consistent with the increased area occupied
by forest in this well preserved region over the study period (; ). In contrast, bird species highly associated with the underbrush (such as Prunella modularis, Sylvia atricapilla, and Troglodytes troglodytes) have increased in the Moreda’s chestnut forest, the montane forest and the holm
oak forest, which could indicate that the shrub substrate has increased in these deciduous
forests. Surprisingly, Phylloscopus ibericus consistently decreased in all the study sites, which might be explained by the relevant
population fluctuations of trans-Saharan migratory species observed (). Lastly, the general declined observed in Erithacus rubecula and Troglodytes troglodytes (excepting at the Moredas’s chestnut woodland), but also the site differences observed
in other species with no changes over time (Fringilla coelebs, Periparus ater, Regulus ignicapilla, Sylvia undata and Turdus merula) are difficult to interpret, partly due to site-specific factors that could be acting
at each site and promoting different dynamics of species population trends.
Finally, the absence of a significant effect of species phenotypic traits on the change
in abundance experienced by the 15 most abundant species can be explained by the fact
that these 15 species did not differ enough from each other (in the phenotypical space
studied with 27 avian traits, which were mainly body size and breeding strategy) to
detect a relationship of temporal patterns in species abundance and bird phenotypic
traits. This result might suggest that different bird traits to that included in our
species phenotypes analysis, could explain specific changes observed in our 15 species
over time, such as the habitat preference or breeding habitat specialization (), but also migration distance (; ) or even biogeographical origin ().
Conclusions
To conclude, our study shows no changes in bird richness and overall bird abundance
after 37 years in a well conserved mountain area of NW Iberia (O Courel) and finds
changes in species specific trends (increasing trends in 5 from the 15 more abundant
species, mainly forest-dwelling species, and decreasing trends in 3 species) which
are unrelated to the species phenotypic traits. Our results indicate that bird communities
in this well conserved region are maintaining their levels of overall diversity, despite
possible site-specific shifts in species abundances. Further investigations in well
preserved areas long-term linked to human presence will be relevant to understand
the rate of change in biodiversity at local scales in the context of a globalized
world.
Acknowledgments
We thank Eduardo J. Castro for designing bird illustrations.
References
1
Allen, D. C., Bateman, H. L., Warren, P. S., Albuquerque, F. S., Arnett‐Romero, S.,
& Harding, B. (2019). Long‐term effects of land‐use change on bird communities depend
on spatial scale and land‐use type. Ecosphere, 10(11). https://doi.org/10.1002/ecs2.2952
2
Bas, S., Guitián, J. M., Sánchez, J. L., & De Castro, A. (1978). Contribución al estudio de las comunidades de vertebrados terrestres de la sierra
de Caurel (Lugo).
3
Bas, S., Guitián, J. M., & Sobral, M. (2018). Biodiversity in Eucalyptus plantations
versus native oak forests in Galicia: plants and birds. Nova Acta Científica Compostelana, 25, 71–81. https://revistas.usc.gal/index.php/nacc/article/view/5192
4
Batáry, P., Kurucz, K., Suarez‐Rubio, M., & Chamberlain, D. E. (2018). Non‐linearities
in bird responses across urbanization gradients: A meta‐analysis. Global Change Biology, 24(3), 1046–1054. https://doi.org/10.1111/gcb.13964
5
Beilin, R., Lindborg, R., Stenseke, M., Pereira, H. M., Llausàs, A., Slätmo, E., Cerqueira,
Y., Navarro, L., Rodrigues, P., Reichelt, N., Munro, N., & Queiroz, C. (2014). Analysing
how drivers of agricultural land abandonment affect biodiversity and cultural landscapes
using case studies from Scandinavia, Iberia and Oceania. Land Use Policy, 36, 60–72. https://doi.org/10.1016/j.landusepol.2013.07.003
6
Blair, R. B. (1999). Birds and butterflies along an urban gradient: Surrogate taxa
for assessing biodiversity? Ecological Applications, 9(1), 164–170. https://doi.org/10.2307/2641176
7
Bowler, D. E., Bjorkman, A. D., Dornelas, M., Myers‐Smith, I. H., Navarro, L. M.,
Niamir, A., Supp, S. R., Waldock, C., Winter, M., Vellend, M., Blowes, S. A., Böhning‐Gaese,
K., Bruelheide, H., Elahi, R., Antão, L. H., Hines, J., Isbell, F., Jones, H. P.,
Magurran, A. E., … Bates, A. E. (2020). Mapping human pressures on biodiversity across
the planet uncovers anthropogenic threat complexes. People and Nature, 2(2), 380–394. https://doi.org/10.1002/pan3.10071
8
Brandt, J. S., Wood, E. M., Pidgeon, A. M., Han, L.-X., Fang, Z., & Radeloff, V. C.
(2013). Sacred forests are keystone structures for forest bird conservation in southwest
China’s Himalayan Mountains. Biological Conservation, 166, 34–42. https://doi.org/10.1016/j.biocon.2013.06.014
9
Buhk, C., Alt, M., Steinbauer, M. J., Beierkuhnlein, C., Warren, S. D., & Jentsch,
A. (2017). Homogenizing and diversifying effects of intensive agricultural land-use
on plant species beta diversity in Central Europe — A call to adapt our conservation
measures. Science of The Total Environment, 576, 225–233. https://doi.org/10.1016/j.scitotenv.2016.10.106
Calviño-Cancela, M. (2013). Effectiveness of eucalypt plantations as a surrogate habitat
for birds. Forest Ecology and Management, 310, 692–699. https://doi.org/10.1016/j.foreco.2013.09.014
12
Coetzee, B. W. T., & Chown, S. L. (2016). Land-use change promotes avian diversity
at the expense of species with unique traits. Ecology and Evolution, 6(21), 7610–7622. https://doi.org/10.1002/ece3.2389
13
Corbelle-Rico, E., Crecente-Maseda, R., & Santé-Riveira, I. (2012). Multi-scale assessment
and spatial modelling of agricultural land abandonment in a European peripheral region:
Galicia (Spain), 1956–2004. Land Use Policy, 29(3), 493–501. https://doi.org/10.1016/j.landusepol.2011.08.008
14
Da Silveira, N. S., Niebuhr, B. B. S., Muylaert, R. de L., Ribeiro, M. C., & Pizo,
M. A. (2016). Effects of land cover on the movement of frugivorous birds in a heterogeneous
landscape. PLOS ONE, 11(6), e0156688. https://doi.org/10.1371/journal.pone.0156688
15
Díaz, L. (2006). Influences of forest type and forest structure on bird communities
in oak and pine woodlands in Spain. Forest Ecology and Management, 223(1–3), 54–65. https://doi.org/10.1016/j.foreco.2005.10.061
16
Donald, P. F., Green, R. E., & Heath, M. F. (2001). Agricultural intensification and
the collapse of Europe’s farmland bird populations. Proceedings of the Royal Society of London. Series B: Biological Sciences, 268(1462), 25–29. https://doi.org/10.1098/rspb.2000.1325
Flohre, A., Fischer, C., Aavik, T., Bengtsson, J., Berendse, F., Bommarco, R., Ceryngier,
P., Clement, L. W., Dennis, C., Eggers, S., Emmerson, M., Geiger, F., Guerrero, I.,
Hawro, V., Inchausti, P., Liira, J., Morales, M. B., Oñate, J. J., Pärt, T., … Tscharntke,
T. (2011). Agricultural intensification and biodiversity partitioning in European
landscapes comparing plants, carabids, and birds. Ecological Applications, 21(5), 1772–1781. https://doi.org/10.1890/10-0645.1
19
Fonderflick, J., Besnard, A., & Martin, J.-L. (2013). Species traits and the response
of open-habitat species to forest edge in landscape mosaics. Oikos, 122(1), 42–51. https://doi.org/10.1111/j.1600-0706.2012.20308.x
20
Fuller, R. J., Gregory, R. D., Gibbons, D. W., Marchant, J. H., Wilson, J. D., Baillie,
S. R., & Carter, N. (1995). Population declines and range contractions among lowland
farmland birds in Britain. Conservation Biology, 9(6), 1425–1441. https://doi.org/10.1046/j.1523-1739.1995.09061425.x
21
Fuller, R. J., Smith, K. W., Grice, P. V., Currie, F. A., & Quine, C. P. (2007). Habitat
change and woodland birds in Britain: implications for management and future research.
Ibis, 149(SUPPL. 2), 261–268. https://doi.org/10.1111/j.1474-919X.2007.00775.x
22
Gao, X., Liang, S., & He, B. (2019). Detected global agricultural greening from satellite
data. Agricultural and Forest Meteorology, 276.277, 107652. https://doi.org/10.1016/j.agrformet.2019.107652
23
García, E., García-Rovés, P., Vigil Morán, A., Alonso, L. M., Fernández Pajuelo, M.
A., Silva, G., Pascual, D., & Álvarez, D. (2014). Atlas de las aves nidificantes de Asturies (1990-2010) (First ed.). Coordinadora Ornitolóxica d’Asturies. https://doi.org/10.13140/2.1.4361.6962
24
Goded, S., Ekroos, J., Domínguez, J., Azcárate, J. G., Guitián, J. A., & Smith, H.
G. (2019). Effects of eucalyptus plantations on avian and herb species richness and
composition in North-West Spain. Global Ecology and Conservation, 19, e00690. https://doi.org/10.1016/j.gecco.2019.e00690
25
Guerrero, I., Morales, M. B., Oñate, J. J., Geiger, F., Berendse, F., Snoo, G. de,
Eggers, S., Pärt, T., Bengtsson, J., Clement, L. W., Weisser, W. W., Olszewski, A.,
Ceryngier, P., Hawro, V., Liira, J., Aavik, T., Fischer, C., Flohre, A., Thies, C.,
& Tscharntke, T. (2012). Response of ground-nesting farmland birds to agricultural
intensification across Europe: Landscape and field level management factors. Biological Conservation, 152, 74–80. https://doi.org/10.1016/j.biocon.2012.04.001
Guitián, J., Guitián, P., Magrach, A., Docampo, C., Domínguez, P., & Guitián, L. (2012).
Effect of management and spatial characteristics on plant species richness of Castanea
sativa Mill. woodlots in the NW Iberian Peninsula. Journal of Forest Research, 17(1), 98–104. https://doi.org/10.1007/s10310-011-0261-x
28
Guitián, J. M., Munilla, I., González, M., & Arias, M. (2004). Guía de las Aves de O Caurel. Lynx Edicions.
29
Herzon, I., Marja, R., Menshikova, S., & Kondratyev, A. (2014). Farmland bird communities
in an agricultural landscape in Northwest Russia: Seasonal and spatial patterns. Agriculture, Ecosystems & Environment, 183, 78–85. https://doi.org/10.1016/j.agee.2013.10.015
30
Järvinen, O., & Väisänen, R. A. (1976). Finnish line transect censuses. Ornis Fennica, 53, 115–118.
31
Jetz, W., Wilcove, D. S., & Dobson, A. P. (2007). Projected impacts of climate and
land-use change on the global diversity of birds. PLoS Biology, 5(6), e157. https://doi.org/10.1371/journal.pbio.0050157
32
Jiguet, F., Devictor, V., Ottvall, R., Van Turnhout, C., Van der Jeugd, H., & Lindström,
Å. (2010). Bird population trends are linearly affected by climate change along species
thermal ranges. Proceedings of the Royal Society B: Biological Sciences, 277(1700), 3601–3608. https://doi.org/10.1098/rspb.2010.0796
Lehikoinen, A., Brotons, L., Calladine, J., Campedelli, T., Escandell, V., Flousek,
J., Grueneberg, C., Haas, F., Harris, S., Herrando, S., Husby, M., Jiguet, F., Kålås,
J. A., Lindström, Å., Lorrillière, R., Molina, B., Pladevall, C., Calvi, G., Sattler,
T., … Trautmann, S. (2019). Declining population trends of European mountain birds.
Global Change Biology, 25(2), 577–588. https://doi.org/10.1111/gcb.14522
35
López-Bao, J. V., González-Varo, J. P., & Guitián, J. M. (2015). Mutualistic relationships
under landscape change: Carnivorous mammals and plants after 30 years of land abandonment.
Basic and Applied Ecology, 16(2), 152–161. https://doi.org/10.1016/j.baae.2014.12.001
36
MacDonald, D., Crabtree, J. ., Wiesinger, G., Dax, T., Stamou, N., Fleury, P., Gutierrez
Lazpita, J., & Gibon, A. (2000). Agricultural abandonment in mountain areas of Europe:
Environmental consequences and policy response. Journal of Environmental Management, 59(1), 47–69. https://doi.org/10.1006/jema.1999.0335
37
McGill, B. J., Dornelas, M., Gotelli, N. J., & Magurran, A. E. (2015). Fifteen forms
of biodiversity trend in the Anthropocene. Trends in Ecology & Evolution, 30(2), 104–113. https://doi.org/10.1016/j.tree.2014.11.006
38
Morelli, F., Benedetti, Y., & Callaghan, C. T. (2020). Ecological specialization and
population trends in European breeding birds. Global Ecology and Conservation, 22, e00996. https://doi.org/10.1016/j.gecco.2020.e00996
39
Munilla, I., López-Bao, J. V., González-Varo, J. P., & Guitián, J. M. (2008). Long-term
changes in the breeding bird assemblages of two woodland patches in Northwest Spain.
Ardeola, 55(2), 221–227. https://www.ardeola.org/es/volumenes/552/articulos/221-227/
40
Parody, J. M., Cuthbert, F. J., & Decker, E. H. (2001). The effect of 50 years of
landscape change on species richness and community composition. Global Ecology and Biogeography, 10(3), 305–313. https://doi.org/10.1046/j.1466-822X.2001.00233.x
41
Payevsky, V. A. (2006). Mechanisms of population dynamics in trans-Saharan migrant
birds: A review. Entomological Review, 86(S1), S82–S94. https://doi.org/10.1134/S001387380610006X
42
Redlich, S., Martin, E. A., Wende, B., & Steffan-Dewenter, I. (2018). Landscape heterogeneity
rather than crop diversity mediates bird diversity in agricultural landscapes. PLOS ONE, 13(8), e0200438. https://doi.org/10.1371/journal.pone.0200438
43
Regos, A., Domínguez, J., Gil-Tena, A., Brotons, L., Ninyerola, M., & Pons, X. (2016).
Rural abandoned landscapes and bird assemblages: winners and losers in the rewilding
of a marginal mountain area (NW Spain). Regional Environmental Change, 16(1), 199–211. https://doi.org/10.1007/s10113-014-0740-7
44
Rosenvald, R., Lõhmus, A., Kraut, A., & Remm, L. (2011). Bird communities in hemiboreal
old-growth forests: The roles of food supply, stand structure, and site type. Forest Ecology and Management, 262(8), 1541–1550. https://doi.org/10.1016/j.foreco.2011.07.002
Salaverri, L., Guitián, J. M., Munilla, I., & Sobral, M. (2019). Bird richness decreases
with the abandonment of agriculture in a rural region of SW Europe. Regional Environmental Change, 19(1), 245–250. https://doi.org/10.1007/s10113-018-1375-x
47
Şekercioğlu, Ç. H., Mendenhall, C. D., Oviedo-Brenes, F., Horns, J. J., Ehrlich, P.
R., & Daily, G. C. (2019). Long-term declines in bird populations in tropical agricultural
countryside. Proceedings of the National Academy of Sciences, 116(20), 9903–9912. https://doi.org/10.1073/pnas.1802732116
48
Seoane, J., Laiolo, P., & Obeso, J. R. (2017). Abundance leads to more species, particularly
in complex habitats: a test of the increased population size hypotheses in bird communities.
Journal of Biogeography, 44(3), 556–566. https://doi.org/10.1111/jbi.12877
49
Shivanna, K. R. (2020). The sixth mass extinction crisis and its impact on biodiversity
and human welfare. Resonance, 25(1), 93–109. https://doi.org/10.1007/s12045-019-0924-z
50
Sirami, C., Brotons, L., Burfield, I., Fonderflick, J., & Martin, J.-L. (2008). Is
land abandonment having an impact on biodiversity? A meta-analytical approach to bird
distribution changes in the north-western Mediterranean. Biological Conservation, 141(2), 450–459. https://doi.org/10.1016/j.biocon.2007.10.015
51
Sirami, C., Brotons, L., & Martin, J.-L. (2007). Vegetation and songbird response
to land abandonment: from landscape to census plot. Diversity and Distributions, 13(1), 42–52. https://doi.org/10.1111/j.1472-4642.2006.00297.x
52
Storchová, L., & Hořák, D. (2018). Life-history characteristics of European birds.
Global Ecology and Biogeography, 27(4), 400–406. https://doi.org/10.1111/geb.12709
53
Suárez-Seoane, S., Osborne, P. E., & Baudry, J. (2002). Responses of birds of different
biogeographic origins and habitat requirements to agricultural land abandonment in
northern Spain. Biological Conservation, 105(3), 333–344. https://doi.org/10.1016/S0006-3207(01)00213-0
54
Tellería, J. L. (2019). Altitudinal shifts in forest birds in a Mediterranean mountain
range: causes and conservation prospects. Bird Conservation International, 30(4), 1–11. https://doi.org/10.1017/S0959270919000455
55
Traba, J., & Morales, M. B. (2019). The decline of farmland birds in Spain is strongly
associated to the loss of fallowland. Scientific Reports, 9(1), 9473. https://doi.org/10.1038/s41598-019-45854-0
56
Valero, E., Álvarez, X., & Picos, J. (2019). Connectivity study in Northwest Spain:
barriers, impedances, and corridors. Sustainability, 11(18), 5124. https://doi.org/10.3390/su11185124
57
Verhulst, J., Báldi, A., & Kleijn, D. (2004). Relationship between land-use intensity
and species richness and abundance of birds in Hungary. Agriculture, Ecosystems & Environment, 104(3), 465–473. https://doi.org/10.1016/j.agee.2004.01.043
Supplementary Materials
Figure S1GLMs results for the variation of (a) species richness (total no. of species) and
(b) abundance (sum of individuals corrected by the nº of surveys) per site and year.
No significant changes were found between years, despite the significant global differences
among sites (a) heathland: p-value<0.010; (b) heathland: p-value<0.001).
Table SISummary of (a) site characteristics (Bas et al. 1978), and (b) bird surveys details
per site (montane forest, holm-oak forest, heathland, Parada’s chestnut woodland,
Moreda’s chestnut woodland) and year (1977, 2014).
Site
Montane forest
Holm oak forest
Heathland
Parada's chestnut woodland
Moreda's chestnut woodland
(a)
Area (ha)
2.50
3.04
3.08
1.80
1.65
Orientation
N
S
S
W
NO-N-NE
Elevation (m.a.s.l.)
1,230-1,320
650
1,000
750-770
760-780
Tree cover (%)
100
70
100
100
90
Tree richness / 400 m2 plot
4
3
0
2
4
Main human influences
Timber harvesting
Firewood collection
Regular fires
Annual chestnut collection + Timber harvesting
Occasional chestnut collection
Surrounding land uses
Oak copses + Heathlands + Mowing fields
Pasturelands + Heathlands + Mowing fields
Rye fields + Oak copses
Croplands and gardens + Mowing fields + Urban
Poor croplands + Meadows
(b)
Year
1977
2014
1977
2014
1977
2014
1977
2014
1977
2014
Length (m)
500
500
530
686
615
615
360
360
330
330
No. of surveys
6
2
3
2
6
2
7
2
8
2
Table SIIPC factor loadings extracted from the PCA for dimensional reduction of avian phenotypic
traits (27 variables with complete cases for the 15 most abundant bird species). The
PC loadings correspond to the three PCs extracted from PCA with eigenvalues >1 and
which explained a total 91.66% of variance in phenotypic traits among bird species.
Phenotypic traits
PC1
PC2
PC3
Bird length - mean (cm)
0.992
-0.051
0.076
Wing length - mean (mm)
0.980
0.012
0.024
Wing length - mean - males (mm)
0.978
0.009
0.026
Wing length - mean - females (mm)
0.981
0.015
0.023
Tail length - mean (mm)
0.959
-0.131
0.168
Tail length - mean of males (mm)
0.958
-0.129
0.169
Tail length - mean of females (mm)
0.960
-0.133
0.168
Bill length - mean (mm)
0.926
0.120
-0.315
Bill length - mean of males (mm)
0.926
0.131
-0.312
Bill length - mean of females (mm)
0.927
0.108
-0.318
Tarsus length - mean (mm)
0.962
-0.063
0.135
Tarsus length - mean - males (mm)
0.963
-0.055
0.147
Tarsus length - mean of females (mm)
0.962
-0.071
0.123
Weight during breeding - mean (g)
0.991
0.002
0.070
Weight during breeding – mean of males (g)
0.991
0.013
0.058
Weight during breeding – mean of females (g)
0.990
-0.009
0.081
Clutch size - minimum (eggs)
-0.277
0.813
0.124
Clutch size - maximum (eggs)
-0.123
0.838
0.402
Clutch size - mean (eggs)
-0.216
0.868
0.395
Brood size per year - mean (broods)
-0.188
-0.522
0.366
Egg length - mean (mm)
0.953
-0.218
0.070
Egg width - mean (mm)
0.955
-0.213
0.074
Egg mass - mean (g)
0.973
-0.143
0.123
Incubation period (days)
0.408
0.717
-0.248
Fledging period (days)
0.367
0.861
-0.068
Feeding independence (days)
0.783
0.460
-0.096
Age of first breeding (years)
0.870
0.266
-0.197
Table SIIIGLM results for the factorial abundance change (increase, decrease and non-significant
change) between years (1977-2014) in function of the three PCs extracted from PCA
(PC1, PC2, PC3) for the phenotype analysis (27 phenotypic traits of the 15 most abundant
bird species), included as covariates (n=15 species). No statistically significant
effects were found.
Response variable
Factors
β
SE
Wald Chi-square
d.f.
P-value
Factorial abundance change
PC1
-0.263
0.579
0.207
1
0.649
PC2
-0.908
0.595
2.325
1
0.127
PC3
-0.490
0.571
0.735
1
0.391
Otros datos estadísticos
Imágenes
María Losada
Me interesan la Biología de la Conservación y la Ecología Evolutiva. Me licencié en Biología en la Universidade de Santiago de Compostela (Galicia), mientras colaboraba con el grupo de investigación de Ecología Evolutiva de Plantas (GI-1287 USC) en el proyecto "Polimorfismo de color y variación geográfica en las interacciones y la selección fenotípica. El caso de Gentiana lutea en la Cordillera Cantábrica" como ayudante de laboratorio y de campo. En 2015, defendí mi tesis de licenciatura centrada en las interacciones de polinizadores y herbívoros con la especie vegetal estudiada, y obtuve el Premio Extraordinario de Licenciatura en Biología. En 2016, cursé el Máster en Biodiversidad y Conservación de la Biología en la Universidad Pablo de Olavide (Sevilla) y colaboré con el Departamento de Ecología Evolutiva de la Estación Biológica de Doñana (EBD-CSIC), defendiendo mi proyecto de fin de máster sobre la asignación del sexo de las crías en una población de papamoscas ibérico (Ficedula hypoleuca) estudiada a largo plazo en La Hiruela (Madrid). Posteriormente, he colaborado como ayudante de campo en otros proyectos de investigación de la Estación Biológica de Doñana (EBD-CSIC), del centro de rescate CERAS (Asociación Quercus, Portugal) y en la consultoría ambiental gallega. Recientemente, he comenzado mi tesis doctoral centrada en el papel ecológico de las interacciones planta-animal en los ciclos biogeoquímicos, en el grupo de investigación EcoPast (GI-1553) de la Universidade de Santiago de Compostela.